OfferGet 10% off on Dissertation, Assignments, Essays, Thesis

Molecular machines, are either natural or artificial molecules which convert chemical energy into mechanical forces and motion. The best understood molecular machines in the field of Molecular Biology are the protein molecules, which catalyze most biological reactions occurring in cells. Protein molecules usually undergo structural changes in order to catalyze the conversion of a substrate into product. Biological molecular machines are used to transport cargo (e.g. kinesin), to power movement (e.g., bacterial flagellar motors), to synthesize proteins (e.g., the ribosome) and many other functions of the cell.
Artificial molecular machines are a major advancement in the field of Materials Science. Artificial molecular machines are the molecular machines synthesized by chemists by mimicking the principles used by the nature in the design biological molecular machines. The principles have been conserved in the nature during the course of evolution. Towards this essay submission project, I would like to prepare an essay on this recent advancement in the field of Materials Science, called the Artificial Molecular Machines. Since, such molecular machines are still under research and development, and have not yet been converted into products for use in our day-to-day life, I will discuss a few examples of molecular machines which have been developed in the lab to describe it’s the extent of progress made in the field. And towards evaluation of the innovation, I will list the number of potential applications where molecular machines could be used.
The foundation for this study was laid centuries ago in 1857 by the Scottish botanist Robert Brown who discovered the motion of tiny particles under the microscope (Brown, 1828; Brown, 1828). Later, this phenomenon called Brownian motion was explained by Einstein (Kinetic theory of matter (Einstein, 1905)) and Perrin (experimental verification of Kinetic theory (Perrin, 1923)).
Some of the fundamental concepts to be learnt from Biological Molecular Machines to aid the development of Artificial Molecular Machines have been described by Erbas-Cakmak, Leigh, McTernan, & Nussbaumer (2015). A few important design points to be remembered are – firstly, Biological molecular machines operate in solution and on surfaces under high viscosity conditions (Viscosity is a measure of the resistance of flow due to internal friction), secondly, the random thermal motion (Brownian Motion) of ions (molecules carrying a net positive or negative charge) is utilized for ion binding, , thirdly, frictional forces become less relevant beyond micrometer scales in viscous medium, fourthly, non-covalent interactions govern the molecular machines equilibrium state (a state of balanced forces), fifthly, bio machines are often made by self-assembly processes (multiple monomers join to form a large polymeric structure), sixthly, compartmentalization or isolation from the environment, is required to allow systems to function away from equilibrium.
In the subsequent paragraphs, example applications and recent advancements / experiments are discussed. In Fig. 1, the stimuli-induced control of conformation about a single-bond in triptycene is shown. It is an example of ‘molecular brake –type machine’ in which the free rotation about the single bond is restricted by cation (positive charge on metal atom) binding (Kelly et al., 1994; Kelly, 2001; Sestelo & Kelly, 2002). The two pyridine groups (a type of chemical group with formula C5H5N) must be coplanar to maximize binding with Hg2+. Therefore, there is a competition between binding to Hg2+ and free rotation, due to which the energy barrier, to allow free rotation about a single bond, increases. Thus creating a molecular switch in which one state (say ON state) is Triptycene-free and the second state (OFF state) is Triptycene-Hg2+ (Hg, chemical notation for Mercury metal) .
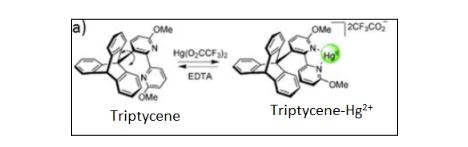
Figure 1 Stimuli-induced control of conformation about a single-bond in triptycene
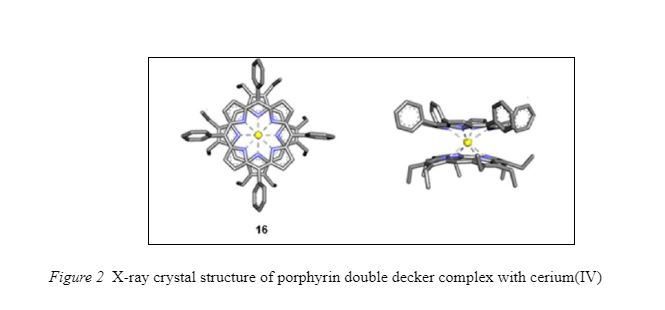
In double-decker organometallic complexes (Tashiro, Konishi, Aida, 1997; Tashiro, Fujiwara, Konishi, Aida, 1998), stimuli induced conformational (structural rearrangement) control may be observed. The rotary motion of ligands in metal sandwich or double-decker complexes is similar to controlled rotation around covalent single bonds. Aida et al. demonstrated that porphyrin ligands in double-decker complexes rotate with respect to each other. The central metal atom and its steric (spatial) bulk influences the speed of rotary motion (Fig. 2).
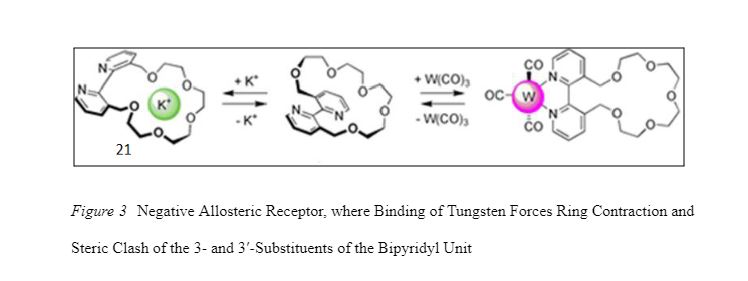
The extraordinary and diverse array of synthetic allosteric receptors (In biochemistry, allosteric regulation (or allosteric control) is the regulation of an protein by binding an effector molecule at a site other than the active site) has gained much interest and attention in recent decades. In biological systems (natural molecular machines), protein-ligand binding is often used to induce conformational change in the host protein. The induced conformational changes in the host could be small, localized, bond deformations or long-range rearrangements. When changes are long-range and across multiple covalent bonds it may cause major conformational change in one of the protein domains with respect to the other. When such conformational changes occur at site in the protein farther away from the ligand-binding site, such a regulation switch is called allosteric regulation. An example of synthetic protein-ligand regulation is the binding of tungsten to allosteric receptor (Rebek, Trend, Wattley& Chakravorti, 1979; Rebek & Wattley, 1980) which reduces the receptor’s affinity for potassium ions (Fig. 3).
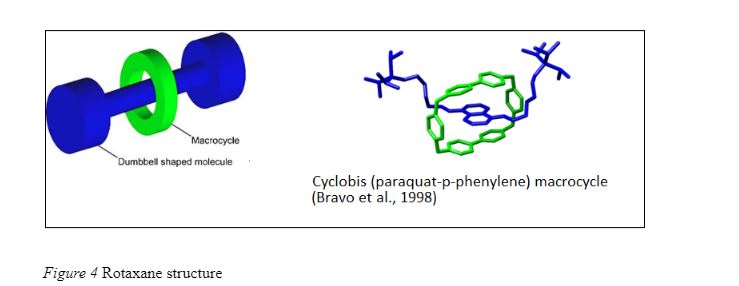
An example of synthetic molecular switches (which work like the electrical switches with separate on/off position) developed by chemists are the Rotaxanes. The name is derived from the Latin word for ‘wheel’ (rota) and axle (axis). A rotaxane has two components – a dumbbell shaped molecule and a macrocycle molecule (Fig. 4, (Bravo, Raymo, Stoddart, White & Williams, 1998)). The two components of a rotaxane remain kinetically trapped because the ends of the dumbbell are larger than the diameter of the macrocycle (large cyclic molecule). In such a trapped structure, the macrocycle is allowed very limited motion in the direction orthogonal to the axle but it is allowed to shuttle along the axis. Due to interaction between the dumbbell and the macrocycle, there are certain distinct sites along the axes at which equillibrim energy state occurs.. Such interactions make rotaxanes ideal molecular machines with multiple ON / OFF states.
DNA (Deoxy Ribonucleic Acid, which code genetic information in cells) macromolecules have been applied to develop molecular machines because of the high specificity of DNA interaction. Many DNA-based molecular motors (Venkataraman, Dirks, Rothemund, Winfree & Pierce, 2007; Tomov et al., 2013), walkers (You et al.,2012 ; Shin & Pierce, 2004), tweezers (Yurke, Turberfield, Mills, Simmel & Neumann 2000), gears (Tian & Mao, 2004), springs (Wang, Huang, Lin, Ren & Qu, 2010), robots (Lund et al.,2010; Muscat, Bath &Turberfield, 2011), transporters (Shelly et al., 2011), and interlocked structures such as DNA rotaxanes and catenanes (Ackermann, Jester, Famulok, 2012; Schmidt & Heckel, 2011) have been developed in the past few decades. Catenanes are mechanically interlocked macrocycles. The fundamental governing principle being sequence-specific interaction based self-assembly of DNA. By this interaction DNA forms double helix and triplex structures. By varying the proportion of nucleotides, for example ATP (Adenosine Tri-Phosphate, a monomer unit in DNA) rich sequence or GTP (Guanosine Tri-Phosphate, a monomer unit in DNA) rich sequences, several ordered assemblies may be generated. The pH induced self-assembly of C-rich sequence into i-motif is an example. The structure of the t-RNA (transfer-RNA) is an example of a natural biological molecular machine. DNA tweezers represent a simple class of DNA machines. DNA tweezers are two-armed contructs bridged by a DNA linker. It can be used to perform molecular motion by response to external stimuli such as pH, temperature, presence of metal ions etc.
To conclude, the advancements made in the past two decades make the future of molecular machines very bright. The 2016 Nobel Prize in Chemistry was awarded to Jean-Pierre Sauvage, Sir J. Fraser Stoddart and Bernard L. Feringa for the synthesis and design of molecular machines. The elaborate research and development studies in this field show a great promise for the future and therefore the research progress may be evaluated to be good and progressing. These synthetic molecular machines have wide potential applications in the future – biomedical diagnostic applications, targeted drug (medicine) delivery for cancer treatment, electronic devices and environmental applications as well. To exemplify its potential use in targeted drug delivery vehicles for chemotherapy / cancer treatment, ‘pH (acidic /basic) responsive molecular machines’ may be designed to take advantage of the very small pH changes between cancerous tissues and normal tissue to release drugs at the targeted cancerous tissue only, thereby promoting safety, fewer side effects to normal tissues. Nanomolecular machines may be utilized in the design of air filters to trap hazardous particulate matter in the air, for pollution control applications. Nanomaterial based molecular machines may also be used in the development of rechargeable batteries and solar cells.
(1) Ackermann, D.; Jester, S.-S.; Famulok, M. (2012). Design Strategy for DNA Rotaxanes with a Mechanically Reinforced PX100 Axle. Angewandte Chemie International Edition, 51, 6771−6775.
(2) Bravo, José A.; Raymo, Françisco M.; Stoddart, J. Fraser; White, Andrew J. P.; Williams, David J. (1998). "High Yielding Template-Directed Syntheses of [2]Rotaxanes". European Journal of Organic Chemistry, 11, 2565–2571.
(3) Brown, R. (1828). A Brief Account of Microscopical Observations Made on the Particles Contained in the Pollen of Plants. Philosophical Magazine, 4, 171−173.
(4) Brown, R. (1828). On the Particles Contained in the Pollen of Plants; and on the General Existence of Active Molecules in Organic and Inorganic Bodies. Edinburgh New Philosophical Journal, 5, 358−371.
(5) Colinvaux, P. (1979). Life at Low Reynolds-Number. Nature, 277, 353−354.
(6) Einstein, A. (1905). Über die von der Molekularkinetischen Theorie der Wärme Geforderte Bewegung von in Ruhenden Flüssigkeiten Suspendierten Teilchen. Annals of Physics, 17, 549−560.
(7). Erbas-Cakmak, S.; Leigh, D.A.; McTernan, C. T.; Nussbaumer, A. L. (2015). Artificial Molecular Machines. Chemical Reviews, 115, 10081 – 10206.
(8) Kelly, T. R. (2001). Progress Toward a Rationally Designed Molecular Motor. Accounts of Chemical Research, 34, 514−522.
(9) Kelly, T. R.; Bowyer, M. C.; Bhaskar, K. V.; Bebbington, D.; Garcia, A.; Lang, F.; Kim, M. H.; Jette, M. P. (1994). A Molecular Brake. Journal of the American Chemical Society, 116, 3657−3658.
(10) Lund, K.; Manzo, A. J.; Dabby, N.; Michelotti, N.; Johnson-Buck, A.; Nangreave, J.; Taylor, S.; Pei, R.; Stojanovic, M. N.; Walter, N.G.; Winfree, E.; Yan, H. (2010). Molecular robots guided by prescriptive landscapes. Nature, 465, 206−210.
(11) Muscat, R. A.; Bath, J.; Turberfield, A. J. (2011). A programmable molecular robot. Nano Letters, 11, 982−987.
(12) Perrin, J. (1923). Atoms (English Translation), 2nd ed. In Hammick, D. L., (Ed.). London:Constable and Co.
(13) Purcell, E. M. (1977). Life at Low Reynolds-Number. American Journal of Physics, 45, 3−11.
(14) Rebek, J.; Trend, J. E.; Wattley, R. V.; Chakravorti, S. (1979). Allosteric Effects in Organic Chemistry. Site-specific Binding. Journal of the American Chemical Society, 101, 4333−4337.
(15) Rebek, J.; Wattley, R. V. (1980). Allosteric Effects. Remote Control of Ion Transport Selectivity. Journal of the American Chemical Society, 102, 4853−4854.
(16) Schmidt, T. L.; Heckel, A. (2011). Construction of a structurally defined double-stranded DNA catenane. Nano Letters, 11, 1739−1742.
(17) Sestelo, J. P.; Kelly, T. R. (2002). A Prototype of a Rationally Designed Chemically Powered Brownian Motor. Applied Physics A: Materials Science & Processing, 75, 337−343.
(18) Shin, J. S.; Pierce, N. A. (2004). A synthetic DNA walker for molecular transport. Journal of the American Chemical Society, 126, 10834−10835.
(19) Tashiro, K.; Konishi, K.; Aida, T. (1997). Enantiomeric Resolution of Chiral Metallobis(porphyrin)s: Studies on Rotatbility of Electronically Coupled Porphyrin Ligands. Angewandte Chemie International Edition, 36, 856−858.
(20) Tashiro, K.; Fujiwara, T.; Konishi, K. (1998). Chemical Communications, 1121−1122.
(21) Tian, Y.; Mao, C. D. (2004). Molecular gears: A pair of DNA circles continuously rolls against each other. Journal of the American Chemical Society, 126, 11410−11411.
(22) Tomov, T. E.; Tsukanov, R.; Liber, M.; Masoud, R.; Plavner, N.; Nir, E. (2013). Rational design of DNA motors: fuel optimization through single-molecule fluorescence. Journal of the American Chemical Society, 135, 11935− 11941.
(23) Venkataraman, S.; Dirks, R. M.; Rothemund, P. W.; Winfree,E.; Pierce, N. A. (2007). An autonomous polymerization motor powered by DNA hybridization. Nature Nanotechnology, 2, 490−494.
(24) Wang, C.; Huang, Z.; Lin, Y.; Ren, J.; Qu, X. (2010). Artificial DNA nano-spring powered by protons. Advanced materials, 22, 2792−2798.
(25) Wickham, Shelley, F. J.; Endo, M.; Katsuda, Y.; Hidaka, K.; Bath, J.; Sugiyama, H.; Turberfield, A. J. (2011). Direct observation of stepwise movement of a synthetic molecular transporter Nature nanotechnology, 6, 166−169.
(26) You, M. X.; Chen, Y.; Zhang, X. B.; Liu, H. P.; Wang, R. W.; Wang, K. L.; Williams, K. R.; Tan, W. H. (2012). An Autonomous and Controllable Light-Driven DNA Walking Device. Angewandte Chemie International Edition, 51, 2457−2460.
(27) Yurke, B.; Turberfield, A. J.; Mills, A. P.; Simmel, F. C.; Neumann, J. L. (2000). A DNA-fueled molecular machine made of DNA. Nature, 406, 605−608.
